摘要:人类基因组计划的宗旨在于揭示人类疾病的遗传根源,并探索更为高效的治疗方法。精神分裂症之所以成为研究的焦点,一方面源于三位项目核心发起人的个人兴趣,另一方面则是基于家庭、领养和双胞胎研究所得出的精神分裂症是遗传性疾病的假设。研究者采取了多种方法进行深入探索,包括连锁分析、候选基因分析、全基因组关联研究(GWAS)、拷贝数变异、外显子组测序等,但并未成功识别出具体的致病基因。不过,研究发现了近300个与精神分裂症发病风险变化相关的单核苷酸多态性(SNPs)以及一些罕见变异,这些在少数个体中可能增加疾病风险。这些风险基因在多数疾病的临床表现中发挥作用,但单独存在时并不足以引发疾病。
随着时间的推移,越来越多的研究人员开始质疑精神分裂症是否严格属于遗传疾病。从1996年起,美国国立精神卫生研究院(NIMH)便开始把研究重点从临床转向基于人类基因组计划的基础研究。因此,该机构三十年来在遗传学方面的投资,几乎未能为受影响的个体带来任何临床上的实用成果。现在,是重新审视NIMH的精神分裂症研究项目的时候了。

▷Torrey,"DidthehumangenomeprojectaffectresearchonSchizophrenia?."PsychiatryResearch333(2024):115691.
本文为文章下篇,主要讲述人类基因组计划三十年来在精神分裂症上做出的努力和结果,并对美国国立精神卫生研究院在项目的方向把握和实施进行反思。
01结果:令人失望的精神分裂症自人类基因组计划伊始,研究人员便利用新兴的技术寻找精神疾病的潜在遗传根源。在过去三十年中,媒体不时宣布关于精神病遗传研究的“重大突破”,但这些所谓的突破往往难以被复制。这一模式——过度的热情紧随其后是失望——与双相情绪障碍的临床表现有着惊人的相似之处。实际上,精神分裂症或许是过去三十年中最频繁被宣布有遗传学突破的疾病。
连锁分析作为90年代广泛采用的一种遗传研究方法,通过新发现的遗传标记来定位与精神疾病相关的染色体区域。这种方法对于有两个或以上家庭成员受影响的家庭特别有效,正如林恩·德里西(LynnDelisi)所识别的那样。其间共有至少32项重大的精神分裂症连锁分析研究(DeLisietal.,2002;Ngetal.,2009)及额外的40项双相情感障碍研究(Kennedyetal.,2015)进行。然而,这些研究的发现都未能得到复制。
2002年,德里西对309个家庭进行了连锁分析,这些家庭至少有两个孩子被诊断为精神分裂症或相关疾病,包括加尔文家庭,其中12个孩子中有7个被诊断为精神分裂症。德里西未能复制先前的连锁研究发现,她的连锁发现同样微弱。她得出结论,连锁分析并非鉴定精神分裂症遗传根源的有效手段(DeLisietal.,2002)。
当连锁分析未能揭示精神疾病的遗传基础时,研究者们转向了候选基因方法,这实质上是一场有教育意义的猜测游戏,试图从约20,000个人类基因中预测哪些可能与疾病相关。例如,鉴于多巴胺在精神分裂症中的作用已知,研究人员将与多巴胺受体相关的基因视为候选基因。到了90年代末和2000年代,有超过1000项精神分裂症候选基因研究进行,估计总成本约为2.5亿美元(Sullivan,2017)。同时,双相情感障碍的候选基因研究也已完成“数百项”(Kennedyetal.,2015)。
遗憾的是,候选基因的研究同样未能取得突破。2015年的一项精神分裂症研究总结指出:“当前的实证证据强烈表明,历史上的候选基因文献未能提供任何有关精神分裂症病因的稳定和可复制的见解”(Farrelletal.,2015)。另一项综述亦得出相似结论:“作为一个整体,精神分裂症候选基因与随机选取的基因组样本无更多关联”(Johnsonetal.,2017)。这些发现促使遗传学领军人物帕特里克·沙利文(PatrickSullivan)建议:“我们应放弃候选基因的猜测……因为它们未带来正确的指引,只是劳而无获”(Sullivan,2017)。
重性抑郁症的候选基因研究也表现不佳。一项涵盖18个候选基因和177,950名抑郁症患者的分析得出结论:“没有确凿证据表明任何候选基因多态性与抑郁症表型有关……总体而言,抑郁症候选基因与非候选基因同样与抑郁症表型无关……我们认为,抑郁症研究应放弃传统的候选基因方法”(Borderetal.,2019)。该研究的首席作者马修·凯勒(MathewKeller)直言不讳地评价:“我们未找到任何证据……我们怎能花费二十年和数亿美元在纯粹的幻觉上?这简直就是《皇帝的新装》的情节,空无一物”(Yong,2019;Marshall,2019)。
连锁分析和候选基因研究未能识别出导致严重精神疾病的任何具体基因,这对于研究者而言无疑是一种挫败。然而,一种新的、充满希望的基因检测方法——广泛称为全基因组关联研究(GWAS)——的出现,为他们提供了新的动力。正如一份报告所标题的那样:“GWAS来拯救”(Offord,2012)。
▷图1GWAS识别的常见等位基因分布图源:/
精神病学中的首项GWAS研究始于2005年。很快,研究者意识到,要获得有意义的结果,需要巨大的样本量。因此,精神病遗传学联盟在2007年成立,旨在鼓励研究者将他们的样本共享至一个公共数据库中,希望这样能够发现一些与精神分裂症原因相关的重要基因。遗憾的是,结果并非如研究者所预期。研究者并没有发现几个与精神分裂症有因果关联的重要基因,反而找到了数百个影响非常微小的基因,这些基因并没有直接因果关系。精神病遗传学联盟最终汇集了76755名患者和243648名对照个体的样本,从中鉴定出了287个增加精神分裂症发病风险的SNP位点(Trubetskoyetal.,2022)。正在进行的双相情感障碍和重性抑郁症GWAS研究涉及更多样本,已鉴定出更多与这两种疾病相关的基因位点。
虽然许多基因位点与增加患病风险的基因相关,但它们也可能起到保护作用,降低某些人群患病的可能性。几乎所有人类疾病都被认为受到这种遗传影响。即便是由感染性病原体引起的疾病,如结核病(由细菌引起)(Phelanetal.,2023;Ghanavietal.,2020)和脊髓灰质炎(由病毒引起)(DeJesus,2007;Pfeiffer,2010),也不例外。人工智能技术的应用,有望通过扩大样本量和检测的SNP数量,进一步探索疾病(包括精神障碍)的遗传联系(Johnson,2023)。
与精神分裂症相关的许多基因位点,位于与疾病有关联但非因果关系的基因附近。例如,已知患有精神分裂症的个体中吸烟成瘾率较高;因此,针对这类个体的GWAS可能会鉴定出与吸烟相关的基因。在精神分裂症个体中鉴定的其他基因位点可能是基于人群差异和病例对照不完全匹配的假阳性。研究者将这一现象称为“隐性人群分层”,并认为它是一个主要的挑战。通过比较欧洲血统和东亚血统个体的抑郁症GWAS研究,突显了该问题的严重性,在欧洲血统个体中鉴定出的基因位点中,仅有11%在东亚群体中也被发现(Giannakopoulouetal.,2021)。
另一个发现是,与精神分裂症相关的大量SNP基因位点也在双相情感障碍、重性抑郁症、自闭症、注意力缺陷多动障碍和焦虑障碍等其他精神疾病的GWAS研究中被报告(Gattetal.,2015;Leggeetal.,2021)。例如,一项研究发现,与精神分裂症相关的基因位点中约50%也与双相情感障碍或自闭症有关(Nestleretal.,2021)。这些结果表明,这些基因位点可能与特定疾病的因果基因无关,而与可能影响多种精神障碍的神经化学路径有关。
其中一项最重要的精神分裂症遗传研究是由林恩·德里西及其同事对特定家庭进行的,这些家庭中多个成员被诊断为精神分裂症或相关精神障碍。研究涵盖了9个家庭,每个家庭至少有3名成员患有精神分裂症,并有其他未受影响家庭成员的DNA可供比对。其中包括加尔文家族,该家族12名成员中有7人被诊断患有精神分裂症。研究者以为,这些负担较重的家庭会帮助他们识别出具有显著影响的致病基因。
然而,在这9个家庭中发现的基因变异并不一致,没有任何一种变异在其他家庭或其他患有精神分裂症的个体中普遍存在。这些发现也没有明确地将变体与疾病原因联系起来,所识别的变异与精神分裂症的GWAS研究中识别的SNP位点没有重叠。例如,在加尔文家族中,所有7名被诊断患有精神分裂症的成员共有一个不同寻常的基因发现,但他们的母亲和一名未患病的姐妹也有同样的基因变异(Hommanetal.,2016;DeLisi,2022b)。然而,其他四名未患病的兄弟姐妹的血液样本不可用,无法确定他们是否也有这种不寻常的基因变异。如果有对精神分裂症有显著影响的致病基因,显然没有在这些负担最重的家庭中找到。
尽管到目前为止全基因组关联研究(GWAS)的成果并不如预期,但不少精神分裂症研究者仍保持着乐观态度,相信这些研究最终会取得突破。他们认为,随着人工智能技术在数据分析中的应用,以及对与已鉴定基因组位点相关的基因功能的进一步理解,我们会有所收获。此外,他们还期待通过鉴定与精神分裂症风险相关的环境因素,GWAS的发现能够帮助揭示基因与环境之间的相互作用。
MHC区域已被研究超过半个世纪,它是一个涉及免疫功能调节的基因区,与许多自身免疫病和传染病有关(Lehner,2012)。1974年,意大利的研究人员首次提出MHC区域与精神分裂症的关联,当时他们怀疑精神分裂症可能是一种自身免疫病(Cazzulloetal.,1974)。随后的研究结果不一,但随着连锁分析方法的出现,MHC在精神病学中的重要性得到了明确。“MHC与精神分裂症的关联是迄今为止在精神遗传学中最为稳定和显著的发现”(NurenbergerandFaroud,1999)。这一发现至今仍然成立,并且如遗传学研究者帕特里克·沙利文所述,它提出了“免疫、自身免疫或感染过程可能对精神分裂症病因的有趣假设”(Sullivan,2010)。
▷GWASdb数据库的人类遗传变异。
尽管有了MHC的重要发现,许多精神分裂症研究界的人士还是对GWAS的成果感到失望。根据最近的一份总结:“针对精神分裂症这一进行了充分赋能的GWAS而言,表现最好的多基因风险评分(PRS)方法只解释了疾病变异的8.5%,当包含非欧洲血统的样本时,这一比例降至7.3%”(Andreassenetal.,2023)。
作者进一步指出,其他精神障碍的GWAS结果“更加落后”。精神病学家乔恩·麦克莱伦(JonMcClellan)和遗传学家玛丽-克莱尔·金(Mary-ClaireKing)指出,GWAS中发现的大多数遗传变异“在疾病的生物学相关性、预后或治疗的临床应用方面尚无明确证据”。随后,他们对进行更多GWAS研究表示怀疑。
“未能证实常见风险变异并非因为GWAS的不当执行。问题在于潜在的生物学,而非研究设计。过去十年,常见疾病-常见变异模型一直是人类基因组学的焦点。已经进行了多项国际合作,研究涵盖数百种重要的人类疾病和特征,涉及大量明显特征的病例和对照组。如果常见等位基因对常见疾病有显著影响,那么现在应该已经发现了许多。问题不在于如何扩大研究规模或进一步分析数据,而在于是否已经对常见疾病-常见变异假说进行了充分测试,并发现它不适用于大多数复杂的人类疾病”(McClellanandKing,2010)。
在连锁分析、候选基因和GWAS研究未能取得预期成果后,精神病学研究者逐渐转向拷贝数变异(CNVs)和外显子组测序,以期发现引发精神分裂症的遗传因素。CNVs是基因组DNA中的常见片段复制或缺失,通常不产生任何不良影响。这些变异大小不一,最大的可涵盖多个基因。一个著名的CNV例子是染色体22区域的大片段缺失,它与DiGeorge综合征或心面综合征有关。这种缺失可能导致多种临床表现,包括心脏、内分泌、泌尿生殖、胃肠、精神和神经问题。发展该综合征的人中就有一个会出现精神分裂症症状。其他可能的表现包括智力障碍、自闭症谱系障碍、注意力缺陷多动障碍、情绪障碍和早发性帕金森病(Bassettetal.,2010;Katoetal.,2023)。
“虽然最初对外显子组测序方法抱有很大的期望,但研究人员逐渐意识到,在精神分裂症的研究中,外显子组测序的诊断效能低下,这些与疾病相关的罕见变异只能在少数精神分裂症患者中发现”(Leeetal.,2023)。即使在这些个体中,这些罕见变异在精神分裂症的发病机制中的作用也不清楚。
外显子组测序和各种类型的拷贝数变异研究仍在继续,技术的不断进步使这些研究得以可能。值得注意的是,迄今为止,几乎所有精神遗传学研究都聚焦在编码蛋白的基因上,这只占我们DNA不到2%的部分。对于剩余的98%的所谓“暗基因组”,即那些没有已知编码蛋白功能的基因组部分,我们知之甚少。最近一项研究筛查了“暗基因组”中的248,000个基因组位点,发现56个在精神分裂症大脑数据中表达有差异(Eradyetal.,2022)。
看来,对于严重精神障碍的遗传成因的探索似乎是一个永无止境的过程。
02讨论:海曼博士的噩梦自比尔·克林顿总统在白宫新闻发布会上与弗朗西斯·柯林斯和克雷格·文特共同宣布完成人类基因组计划以来,已经过去了二十多年。克林顿曾承诺,这项工程将“革改革我们对大多数(如果不是全部)人类疾病的诊断、预防和治疗方式”(Collins,2010)。尤其是在精神疾病领域,弗朗西斯·柯林斯补充道:“精神疾病的诊断和治疗将会发生根本变革”(Collins,2003a)。
然而,二十年后的今天,NIMH在遗传学研究上投入了近80亿美元,却仍未找到任何单一基因导致精神分裂症的证据,也未能带来任何新的治疗方法。在美国,约有400万人患有精神分裂症,包括众多无家可归者和被监禁者,他们的生活状况与人类基因组计划开始前无异。
史蒂文·海曼博士便是这样一位科学家。1986年,他作为分子神经生物学领域的博士后开始研究,并目睹了人类基因组计划的筹备,那时寻找严重精神疾病的致病基因看似充满希望。“我曾认为,家族和遗传学研究、神经科学的进步,加上新兴的分子生物学,很快就能揭示疾病的病理机制,并带来治疗上的突破。但事实证明我错了(Hyman,2018)。”
海曼发现,与其说是单一的因果突变,不如说精神疾病的遗传背景“极其复杂”(Hyman,2011)。他特别对于在有精神疾病家族史的家庭中未能找到致病基因感到失望,“这些家庭中的精神分裂症或躁郁症似乎带有一种悲剧性的必然性”,这里可能指的是琳恩·德里西正在研究的家族。“这些负面结果暴露了遗传复杂性的深层问题,以及一系列令人不安的观察结果(Hyman,2012;Hyman,2018)。”鉴于这些令人失望的成果,海曼在2018年的一次采访中表示担忧,他担心“我们最终只能获得一份基因清单,而不是能推动治疗的机制性理解”(Hyman,2018)。如今,我们确实面对的确实只是一份基因清单。
一些研究者开始质疑,严重精神疾病的所谓因果基因是否真实存在。长期致力于破解精神分裂症遗传谜团的精神病学研究者蒂姆·克劳(TimCrow)于2011年提出了这样的疑问:“当基因数量众多而效果微小时,它们的具体身份几乎变得无关紧要,多基因与无基因之间的界限实际上模糊不清。我们是否已经走到了这一步?”(Crow,2011)
挪威的研究人员也指出:“成千上万的常见遗传变异影响精神障碍”(Andreassonetal.,2023)。精神病学家鲁道夫·乌尔(RudolfUher)和迈克尔·拉特(MichaelRutter)也表示:“总的来说,分子遗传学在精神障碍研究中虽投入巨大,但成果甚微。实际上,在全基因组关联研究的时代,精神障碍在遗传关联的缺失上,与大多数身体疾病形成了鲜明的对比”(UherandRutter,2012)。经过遗传学和病毒学培训的乔纳森·拉瑟姆(JonathanLatham)在其“基因组失败”的文章中也直言不讳:“对常见疾病,如心脏病、癌症和精神疾病的遗传研究中,真正对人类健康有意义的发现寥寥无几。有缺陷的基因很少直接导致疾病,甚至很少让人更倾向于患病……最可能的解释是,这些疾病的遗传基础之所以未被发现,是因为它们几乎不存在”(Latham,2011)。
这三十年寻找精神分裂症致病基因的努力,真的只是人类基因组计划在夸大承诺驱动下盲目的野鹅追逐吗?但事实比这更糟,因为理论上,盲目追逐至少有可能偶然捕获到目标(野鹅)。但如果实际上并不存在导致精神分裂症的单一基因,那么这三十年的探索就更像是在精神病学中寻找不存在的“雪人”。
有两个主要理由让我们怀疑这种探索可能是徒劳的。首先,声称存在导致精神分裂症的基因的证据异常薄弱,这包括对家庭、双胞胎和被领养儿童的研究。
长久以来,精神分裂症在家族中的传播趋势通常被视为其遗传本质的证据。正如一位精神分裂症专家所说,“这无疑是遗传的,它确实在家庭中传播”(Kehres,2020)。普通人患有精神分裂症的终生风险约为1%,但如果一个人的兄弟姐妹患病,该风险升至约10%。虽然这一事实支持了遗传疾病在家族间的传播,但需要注意的是,环境因素——如吸烟、空气污染、含铅油漆、不良饮食习惯和感染——也能以相似的模式在家庭中传播。
其中一个例子是弓形虫病,这种由猫粪中的弓形虫引起的疾病,已知能在某些感染者中触发精神病症状。弓形虫可以通过多种方式感染家庭中的多个成员,例如通过摄入受弓形虫污染的食物或水源(Dubey,2004),或是孩子们在被猫粪污染的沙箱或泥堆中玩耍时感染(Stagnoetal.,1980)。甚至在电脑键盘上也发现了感染性的弓形虫卵囊,表明家庭成员可能通过这种方式感染(Biketal.,2016)。
弓形虫病也可能从怀孕的母亲传给胎儿,这种传播可能在多次怀孕中发生(Asbelletal.,1982)。鉴于人类精子中发现了感染性的卵囊(Diskoetal.,1971),感染的男性很可能将弓形虫传给怀孕的女性,然后再传给她的胎儿。在小鼠中,已记录这种寄生虫可垂直传播多达十代,表现出一种看似遗传的传递模式(Hide,2016)。因此,鉴于这些和其他感染因素在家庭中传播并引发精神症状的多种方式,精神分裂症在家族中的传播不应被视为有力的遗传证据。
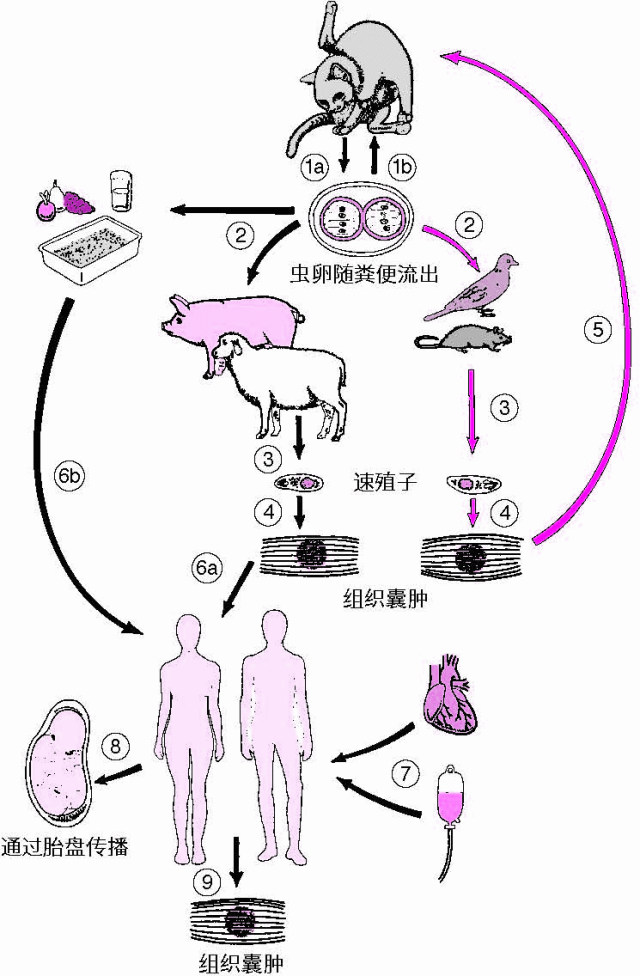
▷图2弓形虫病传染流程图源:默沙东诊疗手册
除了家族研究,从20世纪60年代起,丹麦和芬兰的研究人员通过领养研究来证明精神分裂症的遗传性。主要方法是比较被领养的孩子,区分其生母是否患有精神分裂症。这类研究假设如果生母患病的被领养孩子出现精神分裂症的几率更高,则可作为遗传因素的证据。
我们从家庭研究中知道,患病母亲的子女患精神分裂症的风险约为13%,因此,这些母亲的被领养子女显示出更高的患病风险,并不让人意外(TienariandWynne,1994)。在20世纪60年代和70年代,这一发现被认为具有重大意义,因为当时一些人认为不良的母亲抚养方式是导致精神分裂症的原因。因此,即便是被领养的孩子,在从“不良”母亲转至“良好”母亲的抚养下,仍旧发展出精神分裂症,这一现象被视为对“不良母亲”假设的反驳。
与精神分裂症的家族研究类似,领养研究现在被认为是证明遗传原因相对薄弱的证据。现在我们意识到许多环境因素,包括那些可以影响子宫内发育的感染因素,如母亲分娩时微生物群的传递,以及母乳喂养中传染因素和影响免疫系统的因素也可能从母亲传给新生儿。此外,一些领养研究存在缺陷,因为它们包括了出生数月后才被领养的孩子。所有领养研究在一定程度上存在选择性安置的问题,这一点已被研究人员承认,并认为这种缺陷在某种程度上减弱了遗传因素的影响(Tienari,1992)。
鉴于家庭和领养研究的局限性,研究人员日益依赖双胞胎研究来支持精神分裂症的遗传基础。遗传学家帕特里克·沙利文指出:“我们对精神分裂症病因最坚实且一致的线索是显著的双胞胎/谱系遗传性”(Sullivan,2017)。同卵双胞胎共享100%的基因,而异卵双胞胎只共享50%,与普通兄弟姐妹一样。通过比较同卵和异卵双胞胎患同一疾病的频率,可以估计遗传因素和遗传性的相对重要性。
然而,使用双胞胎评估遗传性存在几个潜在问题,这些问题经常被精神分裂症研究人员忽视。一个问题是双胞胎方法假定同卵和异卵双胞胎的环境因素相等(TenesaandHaley,2013)。实际上并非总是这样。在大约15%的同卵双胞胎中,会存在一个双胞胎在子宫内获得的血液比另一个多,即双胞胎输血综合征,这会导致在母体循环中双胞胎不平等地接触激素、药物和传染因素(Torreyetal.,1996)。双胞胎方法还假定同卵和异卵双胞胎共享相同的社会环境,但实际上,同卵双胞胎相比异卵双胞胎在一起度过更多时间,并拥有更相似的社会网络(Horwitzetal.,2003)。
使用双胞胎研究方法最严肃的问题,早在半个世纪前就被提出了(Rosenthal,1962),即为了确保研究有效,双胞胎样本必须代表整体人群,并且不应基于疾病的严重程度等因素进行选择。然而,用于分析精神分裂症遗传性的多数双胞胎样本实际上是选择性样本。仅有六个未经选择、代表整体人群的双胞胎样本被确定,这些样本全部来自斯堪的纳维亚国家,包括68对同卵双胞胎和654对异卵双胞胎(TorreyandYolken,2019)。选择性样本的问题还因一些精神遗传学家的一种实践而加剧,即如果双胞胎中的每个都被独立诊断,则进行双胞胎对的计算。虽然有研究人员为此辩护(McGue,1992),但其他学者批评这种做法是一种重复计数,人为地夸大了遗传性的估算(Torrey,1992)。
在双胞胎精神疾病研究中的一个关键发现是,在同卵双胞胎对中,两个人都发展成该疾病的比例,即一致率。对于由单个显性基因引起的疾病,如亨廷顿舞蹈症,一致率为100%(14/14)(Sudarskyetal.,1983)。对于患有精神分裂症的同卵双胞胎,一致率在高度选择的样本中,包括重复计数,被报告高达80%,但在现代研究中通常报告为大约50%(Torrey,1992)。
然而,正如TorreyandYolken(2019)展示的,如果一致率仅限于六个未被重复计数的未选择双胞胎样本,则真实的精神分裂症一致率仅为28%(74/268)。实际上,在规模最大且最近的双胞胎研究中,一致率仅为15%(12/81)(Hilkeretal.,2018)。这些较低的一致率与研究感染性疾病的同卵双胞胎研究中发现的一致率相似,例如小儿麻痹症36%(HerndonandJennings,1951),肺结核31%(Bracha,1986),或多发性硬化症27%(Torrey,1992),这些现在被认为是由病毒引起的。这样的一致率被认为反映了对感染剂的共同暴露,而非遗传因素。
总的来说,尽管精神遗传学家通过家族、领养和双胞胎的研究努力证明精神分裂症具有遗传基础,但仔细检查这些证据时,它们支持这一观点的力度出人意料地弱。

▷表1使用基于人群的样本的精神分裂症双胞胎研究
质疑精神分裂症遗传性的第二个主要原因在于,如果存在导致该疾病的基因,那么这些基因必须代代传递以维持疾病的存在。如果疾病影响了个体的生殖能力,则这种疾病将很快消失,因为自然选择非常有效地从种群中移除那些影响生殖健康的遗传变异。
实际上,患有严重精神疾病的人的生育率显著降低。过去半个世纪的研究显示,他们的生育率约为一般人口的三分之一,尽管这一比例随诊断而异。例如,一项来自2010年丹麦的研究报告称,患有精神分裂症的女性生育孩子的可能性仅为未被诊断精神疾病的女性的18%;而患有双相情感障碍的女性为33%,重度抑郁症的女性为57%(LaursenandMunk-Olsen,2010)。
从十九世纪中叶到二十世纪中叶,受严重精神疾病影响的个体的生育率更为低下。在此期间,许多患有这些疾病严重形式的人被长期关押在精神病院中。到了20世纪50年代,美国有超过五十万这样的个体。作为优生学运动的一部分,许多人还被强制接受了绝育手术;在美国,从1907年到1940年,共有18000名精神病患者接受了这种手术。考虑到这极低的繁殖率,人们本可以预期严重精神疾病的患病率会显著下降。然而情况正相反,根据一项研究,从1850年到1950年,在英国的患病率增加了四倍,在美国增加了七倍,在爱尔兰和加拿大增加了八倍(TorreyandMiller,2007)。这一发现与精神疾病遗传起源的假设背道而驰。
纳粹德国进行的一系列行动为这一问题提供了决定性的测试,他们通过限制繁殖以减少精神分裂症患者数量。从1934年到1945年,纳粹绝育了估计有132000名患有精神分裂症的人,并且残忍地杀害了大约同样数量的人。这几乎涵盖了战前德国至少四分之三的精神分裂症患者。战后在德国进行的研究报告了相对较低的现存病例率,正如预期的那样,因为杀害了如此多的患者,但出乎意料的是,新病例数量异常高(TorreyandYolken,2010)。因此,限制精神分裂症患者的繁殖对未来病例数量似乎没有影响,这表明它不太可能是一种遗传疾病。
精神遗传学家长期以来一直被一个谜团所困扰:尽管患有精神分裂症的人群生育率低,这种疾病却意外地持续存在。为了解释这一进化上的悖论,学者们提出了几种可能的假设。
大多数假设认为,受到影响的个体或其直系亲属可能是致病基因的无症状携带者,并且这些基因可能赋予他们某些特殊技能或优势,从而弥补他们在生育健康方面的缺陷。有人提出,患有精神分裂症的人可能更具有创造性。这种权衡的例子是镰状细胞贫血,这种病基因同时也提供了一定程度的抵抗疟疾的能力。然而,目前尚无确凿证据证明,患有精神分裂症或其亲属拥有任何有利特性,能够抵消疾病带来的生育影响(vanDongenandBoomsma,2013)。
另一种解释是,像精神分裂症这样的疾病可能是由于基因组中持续的新变异而产生。遗传学家引用的一个证据是,拥有年长父亲是子女患精神分裂症的一个风险因素,因为年长的父亲更可能传递新的遗传变异。这一点确实成立,但这只能解释不到1%的病例(Torreyetal.,2009)。
总之,对于这一进化悖论,没有令人满意的解释。我们所了解的临床上的精神分裂症已经被详细描述了200多年,在此期间,受影响的人群的生育率极低。如果精神分裂症真是一种遗传性疾病,按理说它现在应该已经消亡了。因此,它的持续存在是一个强有力的证据,表明它并非遗传性疾病。
03结尾:NIMH的问题总结而言,人类基因组计划旨在揭示许多疾病的遗传根源,尤其是精神分裂症及其他严重精神障碍,并寻找更有效的治疗方法。自2000年初步成果公布以来,NIMH已投入约80亿美元于此目标。然而,研究并未找到与精神分裂症有因果关系的单一基因,也未能改善治疗方法。因此,没有确凿证据表明基因缺陷直接导致了精神分裂症。如同绝大多数人类疾病一般,尽管人们发现了与精神分裂症临床表现相关的风险基因,但它们并非疾病的直接原因。
这次研究的失败不仅让400万患有精神分裂症的美国人及其家庭成为了主要的受害者,而且由于NIMH偏重于遗传研究,忽略了其他研究方向,使得其他可能有潜力的研究得不到资助。詹姆斯·塔伯里(JamesTabery)在他的新书《基因的暴政》(TyrannyoftheGene)中指出了一个更广泛的问题,即遗传研究不成比例地主导了多种疾病的研究,但对环境原因的研究可能更有前景。塔伯里认为,这导致了生物医学研究产业偏向于研究健康和疾病遗传原因,不是因为这些因素在健康结果中多么重要,而是因为从事这些研究比研究环境因素更快、更便宜、更有利可图、更符合政治口味(Tabery,2023,)。
NIMH偏爱遗传和基础研究的做法,减少了可能对受影响个体有所帮助的其他研究。例如,通过分析NIMH在2018年资助的与精神分裂症相关的研究项目,我们发现,在428个项目中,仅有7%被认为“可能”在未来20年内为患者带来任何临床改进(Torreyetal.,2020)。临床试验的资金也有所减少,从2006年到2019年,针对精神分裂症的临床试验数量从28项减少到2项,针对躁郁症的从14项减少到0项,针对重度抑郁症的从17项减少到4项(Torreyetal.,2021)。简言之,今天患有精神分裂症的患者的状况并没有比1985年,也就是查尔斯·德里西首次提出人类基因组计划时,有所改善。
那么,接下来我们应该怎么做?回望90年代,人类基因组计划充满希望之时,NIMH对遗传研究的大量投资是合理之举。然而,在过去十年里,遗传研究明显不太可能为精神分裂症提供解答,尽管如此,NIMH仍旧大力投资于此领域,哪怕成果寥寥。NIMH早就应该扩展其研究领域,包括探索不把精神分裂症单纯视作遗传疾病的其他可能。
精神分裂症研究中的一个有前景的方向是研究免疫系统,包括自身免疫疾病、传染病和微生物组失衡等。这一领域之所以充满希望,是因为与主要组织相容性复合体(MHC)在第6号染色体上的相关性是精神分裂症遗传研究中最为稳定的发现之一,而该区域的基因组在遗传上控制着这些功能。遗憾的是,NIMH几乎未投入任何资源来跟进这一发现,可能是因为这一发现并不支持其偏好的精神分裂症遗传性理论。
除了扩展研究领域外,NIMH还需要在基础研究和临床研究之间重新找到平衡。基础研究致力于开发长期的治疗方法,而临床研究旨在为当前遭受痛苦的人们找到更有效的治疗方法。在20世纪90年代之前,NIMH对精神分裂症的基础研究和临床研究保持相对平衡。但自那以后,NIMH在遗传学和其他基础科学研究上的投资不断增加,而临床研究的资金却在减少。原本基础研究和临床研究之间的50/50分配现已变为90/10或更低(Friedman,2015)。这导致了寻找治疗当前受精神分裂症影响人群的更好药物的治疗试验的显著减少。
这些变化应如何在NIMH实施?理论上,它们可以通过NIMH主任或国家精神卫生咨询委员会的命令来实现。然而,当前NIMH主任约书亚·戈登博士(JoshuaGordon)对继续遗传学和其他基础科学研究极其重视。他在2019年的论文中描述的开发更佳精神药物的模型是基于基础研究的(Gordon,2019)。但是,他引用的药物布瑞诺龙(brexanalone),从基础神经化学研究开始到开发完成,花费了35年。对于急需更有效治疗方案的精神分裂症患者来说,更快地获得更有效的药物可能才是更重要的。
国家精神卫生咨询委员会同样不太可能引发NIMH的任何重大改变。理论上,该委员会“向卫生及公共服务部长和NIH院长就进行和支持精神卫生研究的所有政策和活动提供咨询”。实际上,这个由15人组成的委员会,旨在确保所有种族和民族群体、性别、地理区域和精神卫生专业人士都有代表。对重新平衡研究组合感兴趣的委员会成员往往已经获得资助、正在获得资助或希望获得资助,因此不太可能成为改革的积极倡导者。因此,如果要改变NIMH的精神分裂症研究方向,动力很可能需要来自关心此事的国会议员。
人类基因组计划在提升我们对精神分裂症或其他精神障碍的理解和治疗方面未能取得显著进展,但其成败将根据对广泛领域的贡献来衡量。尽管研究对大多数常见疾病的影响有限,但它为受单基因遗传疾病困扰的成千上万家庭带来了治愈的希望。在常见疾病中,癌症或许是从人类基因组计划中受益最大的,因为许多癌症由控制细胞生长的基因突变引起。此外,心理基因组学(psychogenomics)的方法虽充满希望,但至今尚未实现其潜力。人类基因组计划加速发展的技术已改善对传染病病原体的快速测序,为监控流行病如SARS-CoV-2提供了重要工具,同时为CRISPR等基因编辑技术的开发铺平了道路。
此外,人类基因组计划的价值远不止于医学领域。它促进了我们对人类进化的理解,显示出现代智人是数万年前离开非洲的人与尼安德特人、丹尼索瓦人等原始人类的遗传混合体。它还推动了人类基因组与其他232种灵长类动物基因组的比较研究,增进了对人类进化过程的理解(Kudernaetal.,2023)。在法医学领域,改进的测序技术既帮助解决当前犯罪问题,也有助于解开历史悬案。
人类基因组计划的终极益处可能还未完全显现,《经济学人》最近的一篇文章概括道:“因此,基因组学已经成为生物学的基石,如同周期表之于化学。它关联着所有领域。随着这一进展,20世纪80年代的生物学先驱们对他们学科的雄心壮志,正以一种他们未曾预见的方式成为现实”(TheEconomist,2023)。
参考文献:
•Aig,M.,1986.NobelPrizewinner’
•Andreassen,,Hindley,,Frei,O.,Smeland,,2023.Newinsightsfromthelastdecadeofresearchinpsychiatricgenetics,discoveries,(1),4–24..
•Asbell,,Vermund,,Hofeldt,,1982.(5),656–663.(82)90011-3.
•Barry,E.,2022.TheNation’sPsychiatristTakesstock,,2022..
•Bassett,,Scherer,,Brzustowicz,,2010.Copynumbervariationsinschizophrenia,(8),899–914..
•Bassett,,2023.Cli,114992..
•Bik,,Maritz,,Luong,A.,Shin,H.,Dominguez-Bello,,Carlton,,etal.,2016.Microbialcommunityp(6)..
•Birnbaum,R.,Weinberger,,2020.Specialarticle:;canthebridgefrom‘genomics’to‘therapeutics’bedefinedandtraversed?(4),323–329..
•Border,R.,Johnson,,Evans,,Smolen,A.,Berley,N.,Sullivan,,etal.,2019.Nosupportforhistoricalcandidategeneorcandidategene-by-in(5),376–387..
•Bracha,,1986.(12),1634..
•Cazzullo,,1974.Theleuc(0),25–27..
•Collins,F.,2003,CommitteeonEnergyandCommerce,,2003.
•Collins,F.,2003/Fall2003.
•Collins,F.,2010.Hastherevolutionarrived?Nature464(7289),674–675..
•Cook-Deegan,R.,1994.TheGeneWars,Science,Politics,,NewYork.
•Corvin,A.,Morris,,2014.Genome-wideassociationstudies,(4),276–283..
•Costain,G.,Lionel,,Merico,D.,Forsythe,P.,Russell,K.,Lowther,C.,etal.,2013.Pathogenicrarecopynumbervariantsincommunit(22),4485–4501..
•Cowan,,Kandel,,2001.(5),594–600..
•Cowan,,Kopnisky,,Hyman,,2002.,1–50..
•Crow,,2011.Themissinggenes,whathappenedtotheheritabilityofpsychiatricdisorders?(4),362–364..
•DeJesus,,2007.Epidemicstoeradication,,70..
•DeLisi,C.,2022,,Hackensack,NewJersey.
•DeLisi,,Shaw,,Crow,,Shields,G.,Smith,,Larach,V.,etal.,2002.Agenome-widescanforlinkagetochromosomalreg(5),803–812..
•DeLisi,,2022:,22–24..
•Disko,R.,Braveny,I.,Vogel,P.,1971.,391–396.
•Dubey,,2004.(1–2),57–72..
•Erady,C.,Amin,K.,Onilogbo,,Tomasik,J.,Jukes-Jones,R.,Umrania,Y.,etal.,2022.Novelopenreadingframesinhumanacceleratedregionsandtransposableel(3),1455–1468..
•Farrell,,Werge,T.,Sklar,P.,Owen,,Ophoff,,O’Donovan,,etal.,2015.(5),555–562..
•Friedman,R.,2015.“Psychiatry’sIdentityCrisis”.,2015..
•Gatt,,Burton,,Williams,,Schofield,,2015.Specificandcommongenesimplicatedacrossmajormentaldisorders,,1–13..
•Gershon,,Alliey-Rodriguez,N.,Liu,C.,2011.AfterGWAS,(3),253–256..
•Ghanavi,J.,Farnia,P.,Farnia,P.,Velayati,,2020.(3),239–247..
•Giannakopoulou,O.,Lin,K.,Meng,X.,Su,,Kuo,,Peterson,,etal.,2021.Biobankcollaborativegroup,andmajordtry:(11),1258–1269..
•Gordon,,2019.Fromneurobiologytonovelmedications:(6),425–427..
•Herndon,,Jennings,,1951.(1),17–46.
•Hide,G.,2016.Role(3),335–344..
•Hilker,R.,Helenius,D.,Fagerlund,B.,Skytthe,A.,Christensen,K.,Werge,,etal.,2018.Heritabilityofschizophreni(6),492–498..
•Holden,C.,1999..
•Homann,,Misura,K.,Lamas,E.,Sandrock,,Nelson,P.,McDonough,,etal.,2016.Whole-genomesequencinginmultiplexfamilieswithpsychosesr(12),1690–1695..
•Horwitz,,Videon,,Schmitz,,Davis,D.,2003.Rethinkingtwinsandenvironments:(2),111–129.
•Hyman,,2011.(6020),1026..
•Hyman,,2012.(1),3–5.(11)00223-3.
•Hyman,,2018.(1),R6–R8..
•Insel,,Collins,,2003.(4),616–620..
•Isaackson,W.,2021.TheCodeBreaker:JenniferDoudna,GeneEditing,,NewYork.
•Johnson,,Border,R.,Melroy-Greif,,deLeeuw,,Ehringer,,Keller,,2017.Noevidencethatschizophreniacandid(10),702–708..
•Johnson,M.,2023.,2023..
•Kadakia,A.,Catillon,M.,Fan,Q.,Williams,,Marden,,Anderson,A.,2022.(6),22m14458..
•Kato,H.,Catillon,M.,Fan,Q.,Williams,,Marden,,Anderson,A.,etal.,2023.Thegeneticarchitectureofschizophrenia,(3),175–182..
•Kehres,K.,2020.AddressingTabootopics,.
•Keith,,Regier,,Judd,,1988.ReportoftheNationalAdv,1988.
•Kennedy,,Cullen,,DeYoung,,Klimes-Dougan,B.,2015.Thegeneticsofearly-onsetbipolardisorder:,1–12..
•Khullar,D.,2022.,,2022..
•Kolata,G.,1993.ScientistsAtwork,FrancisCollins;,1993;:.
•Kolker,R.,2020.HiddenValleyRoad,,Doubleday.
•Kuderna,,Gao,H.,Janiak,,Kuhlwilm,M.,Orkin,,Bataillon,T.,etal.,2023.(6648),906–913..
•Latham,J.,2011..
•Laursen,,Munk-Olsen,T.,2010.(1–3),234–240..
•Lee,,Ye,R.,Tubbs,,Baum,L.,Zhong,Y.,Leung,,etal.,2023.Third-generationgenomesequencin(11),1058359.
•Legge,,Santoro,,Periyasamy,S.,Okewole,A.,Arsalan,A.,Kowalec,K.,etal.,2021.Geneticarchitectureofschizophrenia:(13),2168–2177..
•Lehner,T.,2012.Thegenesinthemajorhistocompatibilitycomplexasriskfactorsforschizophrenia,(8),615–616..
•Leo,J.,2016.(2)..
•Marshall,L.,2019.Do‘depressiongenes’exist?It’,2019..
•McClellan,J.,King,,2010.(2),210–217..
•McGue,M.,1992.Whenassessingtwinconcordance,(2),171–176..
•Mojtabai,R.,2021.Estimatingthepre,48–49..
•NatlAcademyofScience,1988.,Washington.
•Nestler,E.,Hurd,,Kenny,,Kahn,,,2021.TheHumanBrain,Medicine’:.
•Ng,,Levinson,,Faraone,,Suarez,,DeLisi,,Arinami,T.,etal.,2009.(8),774–785..
•Nurenberger,,Faroud,T.,1998.,79-84.Asquotedin,,,215–218,1999.
•Offord,J.,2012.(2),133–141..
•Pfeiffer,,2010.Innatehostbarrierstoviraltraffickingandpopulationdiversity,,85–118..
•Phelan,J.,Gomez-Gonzalez,,Andreu,N.,Omae,Y.,Toyo-Oka,L.,Yanai,H.,etal.,2023.Genome-widehost-path(1),549..
•Rosenthal,D.,1962.Pro,116–134.
•Schwab,,2022.Geneticvariationandsusceptibilitytoschizophrenia,,114949.
•Shore,D.,ed.,1987.SpecialReport,,USDepartmentofHealthandHumanServices.
•Stagno,S.,Dykes,,Amos,,Head,,Juranek,,Walls,K.,1980.(4),706–712.
•Sudarsky,L.,Myers,,Walshe,,1983.Huntington’(6),408–411..
•Sullivan,,2010.ThepsychiatricGWASconsortium,(2),182–186..
•Sullivan,,2017.Howgoodwerecandidategeneguessesinschizophreniagenetics?(10),696–697..
•Tabery,J.,2023.TyrannyoftheGene,,NewYork.
•Tenesa,A.,Haley,,2013.Theheritabilityofhumandisease,estimation,(2),139–149..
•,2023.HowtheHumanGenomeprojectrevolutionizedbiology..
•Tienari,P.,1992.,52–58.
•Tienari,,Wynne,,1994.(4),233–237..
•Torrey,,1992.Areweoverestimatingthegeneticcontributiontoschizophrenia?(2),159–170..
•Torrey,,Bowler,A.,Taylor,E.,Gottesman,I.,1996.,NewYork.
•Torrey,,Knable,M.,Davis,J.,Gottesman,I.,Flynn,L.,1999.AMissionForgotten:TheFailureoftheNationalInsti,DC..
•Torrey,,2004.,2004.
•Torrey,,Miller,J.,2007.TheInvisiblePlague:,NewBrunswick.
•Torrey,,Buka,S.,Cannon,,Goldstein,,Seidman,,Liu,T.,etal.,2009.Paternalageasariskfactorforschizophrenia,howimportantisit?(1–3),1–5..
•Torrey,,Yolken,,2010.Psychiatricgenocide,(1),26–32..
•Torrey,,Yolken,,2019.Schizophreniaasapseudogeneticdisease:,146–150..
•Torrey,,Knable,,Rush,,Simmons,,Snook,J.,Jaffe,,2020.UsingtheNIHresearch,conditionanddiseasecategorizationdatabaseforresearchadvocacy:(11),e0241062..
•Torrey,,Simmons,,Hancq,,Snook,J.,2021.Theco(11),1342–1344..
•Trubetskoy,V.,Pardiñas,,Qi,T.,Panagiotaropoulou,G.,Awasthi,S.,Bigdeli,,Bryois,J.,etal.,2022.(7906),502–508..
•Uher,R.,Rutter,M.,2012.Basingpsychiatricclassificationonscientificfoundation,(6),591–605..
•vanDongen,J.,Boomsma,,2013.(2),122–136..
•Venter,C.,2007.,Viking.
•Wade,N.,2009.,2009..
•Watson,,1990.TheHumanGenomeProject:past,present,,49–51..
•Yong,,2019.Wasteof1,000,2019..